Tim Lysyk
Agriculture and Agri-Food Canada, Lethbridge, Alberta, CANADA
Contribution No. 3879694
Introduction
Livestock production is an important industry, providing humans with meat, milk, fiber, leather and eggs. The major livestock commodities include beef cattle, dairy cattle, poultry, swine, and sheep. The 1993 USDA Agricultural Statistics Handbook lists the 1992 values of the various commodities as: cattle and calves, $65.5 billion; milk produced by dairy cattle, $20.1 billion; chickens, $9.2 billion for broilers and $3.4 billion for eggs; swine, $4.3 billion; sheep and lambs $0.7 billion.
Arthropod pests of livestock include various species of ticks, mites, lice, fleas, grubs, muscoid flies, mosquitoes, and others. Fortunately, all do not attack livestock at the same time and place. Livestock pest problems vary with season and geographic location, and the pest species exhibit varying degrees of host-specificity and production system-specificity. Integrated pest management (IPM) is the integration of chemical, biological and cultural control methods to reduce pest populations below an economic threshold. Management tools include the conventional methods of monitoring as a basis for control decisions, chemical, cultural and biological controls that individual producers can apply, as well as area-wide controls, sterile male techniques, and regulatory controls that are applied by municipal, provincial/state, or federal governments. Considerable efforts in research, development and implementation have resulted in the elimination of serious threats such as the screwworm, Cochliomyia hominivorax (Coquerel), and cattle tick, Boophilus annulatus (Say), from wide areas of North America (Graham and Hourrigan 1977). However, many problems remain to plague producers at the local level. This discussion will focus mainly on options available to individual producers as area-wide controls involve decisions and techniques beyond the producer's direct control.
Application of livestock pest management practices requires an understanding of the:
- nature of the livestock production system
- biology of the pest arthropod complex associated with the system
- relationship between arthropod populations and damage for a particular system
- extent to which the above influence the ability of the producer to implement chemical, biological and cultural control options
Livestock Production Systems
It is useful to categorize livestock production systems according to their ecological characteristics rather than by commodity, as the characteristics of the system determine the nature of the pest problem and the producer's ability to deal with the problem. Systems are divided into three broad categories (Axtell 1986a):
- grazing (rangeland or pasture)
- outdoor confined
- indoor confined.
Useful descriptions of how various commodities are managed under these systems are available (Martin et al. 1993, Alberta Agriculture 1989).
Grazing systems are characterized by:
- low numbers of animals per unit area
- local vegetation is the main source of animal feed
- management is minimal
- areas are large and manure is dispersed throughout the area
- environment is unpredictable
Grazable vegetation (forage) produced within the system is the main source of animal feed. The amount of forage produced within the system determines the stocking rate, or number of animal units (AU) the system can support. One animal unit (AU) is any number of animals of a species that consumes 11.8 kg of forage per day. A mature 450 kg cow is considered one AU, steers and heifers are 0.67 - 0.75 AU, bulls are 1.5 AU and mature ewes are 0.2 AU. Rangelands are generally unimproved and not suitable for cultivation. Stocking rates are low, down to 0.05 AU per Ha.. Pastures generally have higher stocking rates, up to 2 AU/Ha, and may be fertilized, irrigated and interseeded with improved forages. Cattle are usually turned out to graze in the spring, and are gathered in the summer and branded, dehorned and castrated. Large areas and limited accessibility to animals in grazing systems complicates pest management activities. Manure, the larval habitat of many pestiferous flies, occurs in discreet pats and is widely dispersed. Manure produced by animals fed forages or grasses favors populations of horn flies, Haematobia irritans (L), and face flies, Musca autumnalis De Geer. Off-host populations of many parasites occur within the local vegetation or may develop in standing or flowing water. Many pest species require diapause to successfully overwinter in temperate climates.
Outdoor confined systems are characterized by:
- high numbers of animals per unit area
- concentrated feed is supplied to the system
- management is intensive and animals require daily care
- areas are small and manure accumulates within pens
- some environmental control occurs in sheds and shelters
Outdoor confined systems are those in which animals are contained in pens at relatively high densities. These may be dairies or feedlots for cattle, sheep and swine. Vegetation does not grow within the pen and all feed is provided from external sources. Stocking rates are therefore not limited by carrying capacity of the land area and can be much higher than grazing systems. Feed is usually dispensed onto feed bunks along the front of the pens, and water is provided through a dispensing system. Manure accumulates on the ground and may periodically be pushed into mounds in the centre of the lot. Pestiferous flies breed in the manure mounds and other moist areas. These are in areas behind feed bunks where animals congregate and urinate, or result from leaking waterers, poor drainage or excessive rainfall. Manure produced by animals fed high concentrate feeds favors populations of stable flies, Stomoxys calcitrans (L.), and house flies, Musca domestica (L.). Other breeding sites include spilled feed, stored feed, and undisturbed areas under fences. Increased animal density can promote development of ectoparasite populations, and shelters can increase the duration of ectoparasite problems (Geden et al 1990). Flies overwinter as adults or slowly-developing larvae in sheltered areas.
Indoor confined systems are characterized by:
- extremely high numbers of animals per unit area
- concentrated feed is supplied to the system
- management is intensive and animals require daily care
- areas are small and manure accumulates beneath animals
- buildings are environmentally controlled
Indoor confined systems have the animals housed year-round inside environmentally-controlled buildings. Feed is provided from external sources. Space requirements for animals are minimal and densities can be extreme. High densities favor development of ectoparasite populations and disease outbreaks. Animals are usually supported by expanded steel floors or wire cages and manure accumulates in pits or lagoons beneath. Buildings are environmentally controlled and pest populations may occur year-round, even in cold climates. Numerous resting areas are available for flying insects.
Pest Arthropods and Damage
Pest arthropods in livestock systems can be classified taxonomically or according to their degree of association with the host (Lysyk and Moon 1994). The degree of association with the host is a useful consideration because it determines the management approach. Unfortunately, there is no clear relationship between the degree of association with the host and the relationship between parasite density and production losses. Arthropod pests of livestock can be classified as:
- continuous ectoparasites
- non-continuous ectoparasites
- non-ectoparasites
Continuous ectoparasites complete their entire life-cycle on a host and tend to be host-specific. Continuous ectoparasites include keds, lice, and mites. Non-continuous ectoparasites complete a portion of their life cycle on the host and one or more stages are completed off-host. There are varying degrees of on-host and off-host development. The entire larval stage of cattle grub, Hypoderma spp., develops in the host while pupae and adults occur off-host. Adult horn flies are continually parasitic while the immature stages are free-living. Some species alternate free-living and ectoparasitic phases within a stage. Tick larvae, nymphs and adults all require a blood meal and contact with the host to complete development, but each stage also undergoes some period when not in contact with the host. Adults of many blood-feeding flies feed on the host for short periods and spend much of their time resting off-host to digest the blood meal. Non-ectoparasites, such as the house fly, do not live in direct association with the host but are major problems because they move from animal wastes to nearby human dwellings. Legal actions can be brought against the producer and may result in fines or closure.
Management Thresholds and Economic Injury Levels
Livestock pests can reduce farm productivity by directly reducing animal yields, increasing production costs, or causing missed marketing opportunities. Relationships between density and damage define the goals and needs of livestock IPM. The following types of relationships between pest density and yield-loss apply to livestock IPM:
- presence-absence
- linear
- threshold
- non-linear
The presence of pests on a farm or within a geographic or political area can reduce productivity if the pest vectors disease or is itself classified as a disease. Detection of a single infested animal on a farm may require treatment of all animals if the pest is a quarantinable disease. On a broader scale, import and treatment regulations may be imposed to prevent the introduction of an arthropod or arthropod-borne disease into geographic or political areas. The United States requires that all cattle entering from Mexico be inspected and treated if necessary to prevent introduction of the cattle tick that vectors piroplasmosis. An outbreak of bluetongue disease (vectored by small biting flies in the genus Culicoides) in the Okanagan region of Canada in the 1970's resulted in severe restrictions in semen exports from Canada to European countries because the bluetongue virus can be transmitted through semen as well as by infected vectors. This outbreak had far reaching consequences because importation regulations imposed on cattle entering Canada from the United States increased costs to producers who had to pay for tests to certify that animals were bluetongue free. These import regulations have recently been relaxed.
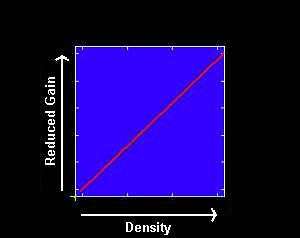
A linear relationship between pest density and production losses can occur. As pest density increases, production losses also increase, either on a per animal or per herd basis. Steelman et al. (1991) estimated that weaning weights of calves decreased by 8% for every 100 horn flies on the cow (averaged over the season) due to reduced milk flow in the cow. Cattle grubs result in a warble that is trimmed from the carcass at slaughter. As the number of warbles increases, the amount of trim loss also increases.
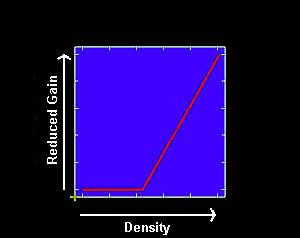
However, density-loss relationships may be linear only for a range of pest densities. Low to moderate populations of many continuous ectoparasites such as lice may have negligible effects on the host (Gibney et al. 1985). Once pest populations reach a threshold density, host defenses may be overcome production losses decrease. Non-ectoparasites such as the house fly may be tolerable at low densities, but at high densities may disperse into neighboring communities, annoying the residents and resulting in legal actions.
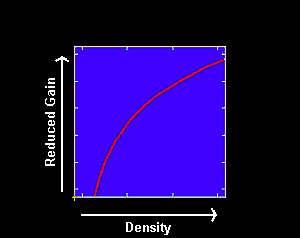
Lone star ticks Amblyomma americanum (L.) show a non-linear relationship between pest density and production losses (Barnard 1985.), with damage approaching an upper asymptote as density increases. Similarly, losses due to horn flies feeding on growing cattle may also be non-linear (Kinzer et al. 1984).
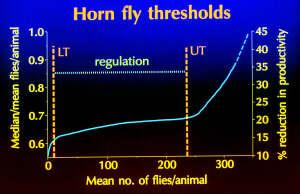
Haufe (1986) proposed a two-threshold model that incorporates aspects of the linear, threshold and non-linear relationships to pest density. Haufe hypothesized that reductions in animal productivity occur at two threshold pest densities. The most obvious threshold is associated with high infestations. The animals' ability to tolerate infestations above this upper threshold is impaired and reductions in productivity increase in a steep linear response to increasing pest density. The lower threshold is a quantal response to low infestation levels and results from behavioral changes that reduce forage utilization. The result is an immediate reduction in productivity that persists until the upper threshold is reached. Management actions can likely be applied to reduce the ability of the pest population to reach the upper threshold, but it may be much more difficult to reduce the pest's ability to reach the lower threshold. Maintenance of the pest population density below the lower threshold may ultimately provide the most consistent and highest economic benefits.
Management in Grazing Systems
Two approaches can be used to monitor pest populations on grazing animals. Visual estimates of the number of ectoparasites per host can be made by counting the number attacking during a fixed time interval. This approach can work with ectoparasites that have prolonged contact with the host, e.g. adult horn flies and ticks. However, visual estimation in grazing systems is complicated by several factors. Direct access to animals is not always possible because the animals generally graze over large areas. Most ectoparasites are small and cannot be readily enumerated. Non-continuous ectoparasites have brief association with the host and not all of the ectoparasite population may be on the host at any one time. An alternate approach is to use traps in fixed locations to determine relative abundance of the free-living stages of an ectoparasite. These relative indices can be used to estimate attack rates if the relationship between the relative estimate and numbers attacking the host are known. Such relationships have been demonstrated for face flies (Moon and Kaya 1981) and mosquitoes (Steelman et al. 1972).
Drummond et al (1988) provide an excellent summary of chemical control methods for most arthropod pests of livestock. Chemical control methods are the management options most frequently used by producers in grazing systems. The animals themselves are generally the target of chemical applications since they are the commodity to be protected and will bear the ectoparasite population at some point in time. Chemical application methods most commonly used are:
- direct application to the animal by spraying or dipping.
- self-application devices such as dust bags or oilers
- sustained release devices such as ear tags and boluses.
Choice of an application method depends on the length of the attack period of the ectoparasite and the ease with which it is applied. Direct treatment by spraying or dipping can provide effective control if the residual activity of the chemical is long relative to the attack period of the ectoparasite. For example, control of tick paralysis in British Columbia requires about 6 - 8 weeks of residual chemical activity if the cattle are treated before being released to pasture. Direct sprays can also provide 1 -2 weeks control of other ectoparasites such as mosquitoes and black flies, but the attack periods of these pests may last over one month. In such cases, reapplication may be necessary, and this requires gathering the animals. Monitoring is essential to determine the need for retreatment.
A major limitation with direct application to animals is the producer's need to gather and confine the animals before treatment. Self-treatment devices that dispense insecticides to animals are often used because these can be placed on pasture and are visited by the animals. Oilers consist of fabric-wrapped metal chains inserted in a reservoir containing insecticide. The fabric wicks the insecticide from the reservoir and the insecticide is dispensed on animals as they rub against the chains. Dust bags contain insecticide dust and are suspended above gates so that the animals contact the bag as they walk through the gate. Because self-applicating devices rely on animal behavior to dispense insecticide, they must be positioned in areas near watering site or salt licks that animals visit frequently. They must also be serviced frequently to ensure that pesticide is present and the wicks are working properly.
Sustained release devices consist of insecticides placed within a matrix. The pesticide is slowly released during the grazing season. Insecticidal ear tags are the most commonly used sustained release device. These consist of insecticides embedded in a plastic matrix. After application, the insecticides diffuse over the oil-soluble surface of cattle. These can be applied before animals are turned out to pasture and can provide season long-control of horn flies because horn flies spend most of their adult life on the host and are exposed to toxic doses of insecticides. They do not control non-continuous ectoparasites such as mosquitoes, black flies, and tabanids. Advantages of ear tags include ease of application, long duration of efficacy, and much smaller amounts of pesticides are placed in the environment and only in specific targets. The major disadvantage of ear tags is that resistant populations of horn flies developed shortly after tags were released. Ear tags were released in the late-1970's, and resistance had become widespread throughout the U.S. by the mid 1980's (Kunz and Schmidt 1985), and throughout Canada by 1991 (Colwell et al. 1991, Mwangala and Galloway 1993).
Factors contributing to the rapid development of resistance include widespread tagging using a single chemical family, the pyrethroids, against entire adult populations of horn flies. Resistance in horn flies is controlled by a set of alleles that express incomplete dominance (Roush et. al 1986). Heterozygous resistant (RS) individuals have fitness between homozygous resistant (RR) and homozygous susceptible (SS) individuals. As the concentration of pesticide released from ear tags decreases throughout the growing season, RS individuals have increased chance of survival compared with SS individuals and the R allele becomes rapidly established in the population. Resistance management has become an important issue for producers and can be achieved by altering application methods from season to the next, and alternating compounds from different chemical classes such as OP and pyrethroid insecticides. Ear tag use should avoid low concentrations of the pesticide that may favor development of RS individuals. Care should be taken not to tag before the onset of the fly season, tags should be applied to target animals (Bulls, steers, cows and not calves) at the maximum recommended rate, and tags should be removed at the end of the fly season or when populations decline before winter.
Sustained-release boluses are currently being marketed. These are blocks that are inserted into the animal's stomach and release insecticides that are absorbed into the animal's bloodstream. Blood-feeding arthropods ingest the insecticide with the blood meal and are killed. The insecticide is also released into the gut and ultimately into the feces. The insecticide in the feces is toxic to developing filth flies such as the horn fly. The insecticides may also be toxic to the beneficial dung arthropods that degrade pats and play an important role in nutrient recycling within pastures and act as natural enemies of the pest flies (Floate, 1996).
Naturally occurring biological control agents can considerably reduce fly production (Doube 1986). Biological control of manure-breeding pasture flies has largely been attempted through establishment of imported natural enemies as predators, parasites, pathogens, and competitors. Numerous species of exotic dung beetles have been imported into the United States and Australia to facilitate dung burial and pat breakdown, thereby reducing the breeding habitat for filth flies. Individual producers cannot undertake such classical biological control efforts, but can use conservation principles to avoid interfering with the action of the natural enemies where they are abundant. Inundative approaches to biological control have not been as vigorously pursued.
Cultural management options have been developed, or are being developed, for a variety of pasture pests. These are perhaps most-well developed for the single-host ticks in the genus Boophilus in Australia and includes use of resistant crosses of cattle, pasture spelling and acaricide treatment (Norton et al. 1984). European cattle breeds are much more susceptible to tick-induced losses than Zebu animals. Unfortunately, Zebu animals have low fertility and slow growth. Traditionally, cattle ticks were controlled on European animals using frequent, well-timed dipping regimes, but this led to acaricide resistance. Profitability can be improved by using cross-bred animals that have moderate growth and are resistant to ticks, and require fewer dippings for effective control, thereby reducing the probability of developing resistance. Use of cross-bred cattle has also led to feasible pasture-spelling strategies. Boophilus ticks complete the larval, nymphal and adult stages on the same host. The adults drop from the host, lay eggs in the soil, and hatched larvae reattach to a new host. Cattle can be confined to a pasture during the period when adults are dropping and removed during larval hatch so that the ticks cannot reinfest new hosts. This strategy was somewhat unfavorable for European breeds and led to overgrazing and undergrazing as pastures could be overstocked during tick drop and underused during larval activity. However, use of resistant crossbred cattle can provide effective tick management with as little as four weeks spelling, a dramatic improvement.
Cultural methods are most highly developed with cattle tick, but host resistance has been identified against horn flies (Steelman et al. 1994). Long-haired cattle also are resistant to black fly attack. The impact of controlled grazing strategies on filth fly production is currently under investigation (R. D. Moon, pers. comm.). Walk through traps that brush flies from cattle are also showing promise in specialized situations (Tozer and Sutherst 1996).
Outdoor Confined Systems
Pest populations can be more readily assessed in outdoor confined systems because producers have direct access to the animals, or can place traps in convenient locations. Stable fly populations are assessed using counts of the number of flies landing on the front legs of animals, or by determining the proportion of infested legs. Small continuous ectoparasites can be monitored by rating infestations according to pest density or severity of symptoms. Non-ectoparasite populations can be monitored using traps. Information from monitoring programs provides the basis for chemical control treatments. Non-ectoparasites such as the house fly do not cause direct losses in animal productivity, but have a high nuisance value as they disperse to nearby residences. Producers and local residents must work together to determine at what on-farm density flies become a problem to nearby residents. This level can be used as an action threshold.
Chemical pesticides are applied using a variety of methods, and applications are more convenient than in grazing systems. Continuous ectoparasites can be managed using pour-on applications of small doses of pesticides applied directly to the animals. Sprays can be readily applied as animals can be rounded up easily. Non-continuous ectoparasites such as adult stable flies and non-ectoparasites such as adult house flies rest on surfaces and can be effectively controlled using residual sprays applied to resting areas. Resting areas can be readily identified by the presence of fecal or vomit spots. Baits can also be applied for adult non-ectoparasites. Pesticide resistance has developed due to overuse of pesticides and can be avoided by monitoring populations, treating only when needed and not on a calendar basis, and rotating chemical families (MacDonald et al. 1984). Direct applications to animals are usually not effective for control of non-continuous ectoparasites and non-ectoparasites as these do not visit the host long enough to pick up a toxic dose.
Chemical control methods are most effective against continuous ectoparasites, adult non-continuous ectoparasites and adult non-ectoparasites. Immature stages of non-continuous ectoparasites and non-ectoparasites usually occur in moist, decaying organic matter such as manure, spilled feed and moist soil. Chemical applications against these stages are not recommended because the breeding media interferes with pesticide contact, moistens the media, and can interfere with the activity of naturally occurring biological control agents such as predaceous mites.
The most commonly used biological control method is inundative releases of small parasitic wasps in the family Pteromalidae. The wasps oviposit inside fly puparia and the developing wasp larvae feed on and kill the fly pupa. The parasites are available from commercial producers. Successful control using these agents alone has been variable, but weekly releases of 200 parasitized pupae (Muscidifurax raptor Girault and Sanders) per cow and 1000 parasitized pupae per calf was an essential component of IPM programs in dairies in New York State. (Rutz and Watson In Press). Pesticides such as pyrethrin space sprays and scatter baits are more compatible with parasitoids than are pesticides from other families or formulations.
Sanitation is required to reduce breeding success of filth flies. Stable fly populations in Nebraska feedlots have been reduced by up to 50% under rigorous sanitation regimes that included complete cleanout of lots before peak stable fly activity followed by biweekly cleanings (Thomas et al. 1996). Sanitation practices used in dairies in New York State required weekly removal and spreading of manure (Rutz and Watson, In Press). Cleaning requires removal of all spilled feeds, manure, silage in and the lots and feed storage areas. wastes can either be spread directly on surrounding fields or stored under plastic to reduce fly breeding potential. Elimination of moisture resulting from improper drainage and leaking waterers is also required. Costs associated with sanitation can be compensated for by reduced pesticide cost (Lazarus et al. 1989).
Management in Indoor Confined Systems
Many of the same practices used for pest management in outdoor confined systems are applicable to indoor confined systems. Confined systems have the added complications of structural pests such as lesser mealworm, but these will not be dealt with here. The major difference between indoor and outdoor confined systems are that arthropod populations may be active year-round in indoor systems where they are protected from the vagaries of weather. Natural enemy populations may also be present year-round and can be augmented using inundative releases of parasitic wasps. Monitoring ectoparasite populations is relatively easy due to ready access to animals. Monitoring non-ectoparasitic pests is also relatively easily accomplished using relative indices of abundance.
Chemical application methods depend on the type of pest involved. Direct spraying of animals is used for continuous ectoparasites. Residual spraying or space sprays are used for free-living pests or non-continuous ectoparasites. Baits are also used against adult flies. Larviciding for control of the immature stages of filth flies is not recommended as larvicides reduce natural enemies in the manure and require considerably more applications than integrated control using selective adulticides that do not interfere with natural enemy populations. (Axtell 1970a). Integrated control using a combination of selective adulticides and conservation of natural enemies provides the same degree of control as frequent larviciding but uses only 20% of the insecticide used for larviciding (Axtell 1970b). Non-continuous ectoparasites can live in tiny cracks and sheltered areas that are difficult to treat. Care must be taken to avoid contamination of feed and watering equipment. Also, frequent use of a single family of pesticides should be avoided to prevent development of resistance.
Biological control is generally not used for ectoparasites but is effective against non-ectoparasites such as filth flies. Biological control agents can occur naturally and their populations can be promoted using conservation principles and manure management. A number of biological control agents are commercially available. The most frequently used agents are pupal parasites in the family Pteromalidae. These provide more reasonable control in indoor confined systems (Rutz and Axtell 1979, Costello 1984, Weintraub 1985) compared with outdoor systems (Peterson et al. 1983, Meyer et al 1990). Augmentation of natural enemies should be considered if manure is managed favorably, the species is not already present, and selective insecticides are being used (Axtell and Rutz 1986).
Sanitation practices for ectoparasite management are required to reduce the possibility these pests may move among houses. Many ectoparasites can live off the host and are easily transmitted from one house to another by contaminated equipment and people, and birds. Cleaning carts, crates and other materials moved from house to house is required. Use of disposable outer clothing by workers can help reduce movement of ectoparasites between houses. Wild birds can spread poultry ectoparasites among houses, but can be excluded from entering the houses by screening. Axtell and Arends (1990) recommend that detection of a single ectoparasite is sufficient cause to treat due to their potential to increase in number and their ability to disperse undetected. A cautious approach is to treat immediately to avoid any risk of contamination of other houses.
Manure management is essential for filth fly control. Many systems allow the manure to accumulate under the cages or floors for lengthy periods, and it is in these that filth flies may become major pests. Substantial populations of natural enemies such as predaceous mites and beetles can develop in dry manure (Axtell 1986b) and provide effective control of fly populations. Natural enemy populations can be encouraged if manure management is conducted with conservation in mind (Axtell 1970a). Eliminating leaking waterers, promoting air circulation to dry manure and providing adequate drainage can reduce manure moisture, inhibit fly larval development, and promote predation by natural enemies (Axtell 1986b). Complete manure removal eliminates the natural enemy fauna and fly populations can quickly rebound after manure removal. Manure removal should therefore be conducted early in the season when fly populations are lowest, and removal should be staggered to allow natural enemies to recolonize disturbed areas.
References
- Alberta Agriculture. 1989. The beef cow-calf manual. Alberta Agriculture, Edmonton, Alberta.
- Axtell, R. C. 1970a. Integrated fly control program for caged-poultry houses. J. Econ. Entomol. 63: 400-405.
- Axtell, R. C. 1970b. Fly control in caged-poultry houses: Comparison of larviciding and integrated control programs. J. Econ. Entomol. 63: 1734-1737.
- Axtell, R. C. 1986. Status and potential of biological control agents in livestock and poultry pest management systems. Entomol. Soc. Am. Misc. Publ. 61: 1-9.
- Axtell, R. C. 1986. Fly management in poultry production: cultural, biological, and chemical. Poultry Science 65: 657-667.
- Axtell, R. C. and J. J. Arends. 1990. Ecology and management of arthropod pests of poultry. Ann. Rev. Entomol. 35: 101-126.
- Axtell, R. C. and D. A. Rutz. 1986. Role of parasites and predators as biological fly control agents in poultry production facilities. Entomol. Soc. Am. Misc. Publ. 61: 88-100.
- Barnard, D. R. 1985. Injury thresholds and production loss functions for the lone star tick, Amblyomma americanum (Acari: Ixodidae), on pastured, preweaner beef cattle, Bos taurus. J. Econ. Entomol. 78: 852-855.
- Campbell, J. B., I. L. Berry, D. J. Boxler, R. L. Davis, D. C. Clanton and G. H. Deutscher. 1987. Effects of stable flies (Diptera: Muscidae) on weight gain and feed efficiency of feedlot cattle. J. Econ. Entomol. 80: 117-119.
- Colwell, D. D., T. J. Lysyk, A. Whiting and H. Philip. 1992. Horn fly resistance widespread. LRS weekly letter No.3056.
- Costello, R. A. 1984. In J. S. Kelleher and M. A. Hulme (Eds.) Biological control programmes against insects and weeds in Canada 1969-1980. Commonwealth Agricultural Bureaux, London. Musca domestica L., House fly (Diptera: Muscidae). pp.63-64
- Doube M. 1986. Biological control of the buffalo fly in Australia: the potential of the South African dung fauna. Misc. Publ. Entomol. Soc. Am. 61: 10-34.
- Drummond, R. O., George J. E., Kunz S. E. 1988. Control of arthropod pests of livestock: a review of technology. CRC Press, Boca Raton, Fla. 245 pp.
- Floate, K. D. 1996. Ivermectin residues in cattle dung: effects on non-target organisms and dung breakdown. Alberta Agricultural Research Institute Final Report No. 940542. 62 pp.
- Geden, C. J., D. A. Rutz and D. R. Bishop. 1990. Cattle lice (Anoplura, Mallophaga) in New York: seasonal population changes, effects of housing type on infestations of calves, and sampling efficiency. J. Econ. Entomol. 83: 1435-1438.
- Gibney, V. J.,J. B. Campbell, D. J. Boxler, D. C. Clanton and G. H. Deutscher. 1985. Effects of various infestation levels of Cattle Lice (Mallophaga: Trichodectidae and Anoplura: Haematopinidae) on feed efficiency and weight gains of beef heifers. J. Econ. Entomol. 78: 1304-1307.
- Graham, O. H. and J. L. Hourrigan. 1977. Eradication programs for the arthropod parasites of livestock. J. Med. Entomol. 13: 629-658
- Haufe, W.O. 1986. A modeling system for horn flies on cattle. Modeling and simulation: tools for management of veterinary pests. USDA ARS. ARS-46. In: Miller, J. A. (Ed.). 1986.
- Kinzer, H. G., W. E. Houghton, J. M. Reeves, S. E. Kunz, J. W. Wallace and N. S. Urquhart. 1984, Influence of horn flies on weight loss in cattle, with notes on prevention of loss by insecticide treatment. Southwest. Entomol. 9: 212-217.
- Kunz, S. E. and C. D. Schmidt. 1985. The pyrethroid resistance problem in the horn fly. J. Agric. Entomol. 2: 358-363.
- Lazarus, W. F., D. A. Rutz, R. W. Miller and D. A. Brown. 1989. Costs of recommended manure management practices for house fly and stable fly (Diptera: Muscidae) control on dairy farms. J. Econ. Entomol. 82: 1145-1151.
- Lysyk, T.J. and R. D. Moon. 1994. Sampling arthropods in livestock management systems. pp. 515 - 538 In: Pedigo, L and D. Buntin (eds.) Handbook of sampling methods for arthropod pests in agriculture, CRC press, Boca Raton, Fla.
- MacDonald, R. S., G. A. Surgeoner, K. R. Solomon and C. R. Harris. 1983. Development of resistance to permethrin and dichlorvos by the house fly (Diptera: Muscidae) following continuous and alternating insecticide use on four farms. Can. Entomol. 115: 1555-1561.
- Martin, J., R. J. Hudson and B. A. Young (Eds.). 1993. Animal production in Canada. University of Alberta Press, Edmonton, Alberta. 340 pp.
- Meyer, J. A., B. A. Mullens, T. L. Cyr and C. Stokes. 1990. Commercial and naturally occurring fly parasitoids (Hymenoptera: Pteromalidae) as biological control agents of stable flies and house flies (Diptera: Muscidae) on California dairies. J. Econ. Entomol. 83: 799-806.
- Moon, R. D. and H. K. Kaya. 1981. A comparison of methods for assessing age structure and abundance in populations of nondiapausing female Musca autumnalis (Diptera: Muscidae). J. Med. Entomol. 18: 289-297.
- Mwangala, F.S. and T. D. Galloway. 1993. Susceptibility of horn flies, Haematobia irritans (L.) (Diptera: Muscidae), to pyrethroids in Manitoba. Can. Entomol. 125: 47-53.
- Norton, G. A., R.W. Sutherst and G. F. Maywald. 1984. Implementation models: the case of the Australian cattle tick. pp. 381-394 In: Conway, G.R. (Ed.) Pest and pathogen control: Strategic, tactical and policy models. John Wiley and Sons, New York.
- Petersen, J. J., J. A. Meyer, D. A. Stage and P. B. Morgan. 1983. Evaluation of sequential releases of Spalangia endius (Hymenoptera: Pteromalidae) for the control of house flies and stable flies (Diptera: Muscidae) associated with confined livestock in eastern Nebraska. J. Econ. Entomol. 76: 283-286.
- Roush, R. T., R. L. Combs, T. C. Randolph, J. MacDonald and J. A. Hawkins. 1986. Inheritance and effective dominance of pyrethroid resistance in the horn fly (Diptera: Muscidae). J. Econ. Entomol. 79: 1178-1182.
- Rutz, D. A. and R. C. Axtell. 1979. Sustained releases of Muscidifurax raptor (Hymenoptera: Pteromalidae) for house fly Musca domestica control in two types of caged-layer poultry houses. Environ. Entomol. 8: 1105-1110.
- Rutz, D.A and D. W. Watson. Parasitoids as a component in an integrated fly management program on dairy farms. In: Ridgeway, R. L., C. S. Glenister and M. Inscoe (Eds.). Mass reared natural enemies: Applications and regulations. Entomological Society of America Monograph, Lanham, MD. In Press.
- Steelman, C. D., A. H. Brown, E. E. Gbur and G. Tolley. 1991. Interactive response of the horn fly (Diptera: Muscidae) and selected breeds of beef cattle. J. Econ. Entomol. 84: 1275-1282.
- Steelman, C. D., R. W. McNew, M. A. Brown, G. Tolley and J. M. Phillips. 1994. Efficacy of Brahman breeding in the management of insecticide-resistant horn flies (Diptera: Muscidae) on beef cattle. J. Econ. Entomol. 87: 7-14.
- Steelman, C. D.,T. W. White and P. E. Schilling. 1972. Effects of mosquitoes on the average daily gain of feedlot steers in southern Louisiana. J. Econ. Entomol. 65: 462-466.
- Thomas, G. D., S. R. Skoda, D. R. Berkebile and J. B. Campbell. 1996. Scheduled sanitation to reduce stable fly (Diptera: Muscidae) populations in beef cattle feedlots. J. Econ. Entomol. 89: 411-414.
- Tozer, R. S. and R. W. Sutherst. 1996. Control of horn fly (Diptera: Muscidae) in Florida with an Australian trap. J. Econ. Entomol. 89: 415-420.
- Weintraub, J. 1985. Biological control of house flies in confined livestock rearing. In L. J. L. Sears and E. E. Swierstra (Eds.) Research Highlights - 1984. Agriculture Canada Research Station.Lethbridge.pp.26-28.