Timothy C. Lockley
Imported Fire Ant Station
USDA/APHIS/PPQ
Gulfport, MS 39501
History

Four species of fire ants are currently found within the contiguous southeastern United States. The tropical fire ant, Solenopsis geminata Fabricius, and the southern fire ant, S. xyloni McCook, are considered species "native" to the area. The two imported species of fire ants were introduced into the United States from South America at the port of Mobile, Alabama. The black imported fire ant, Solenopsis richteri Forel, arrived sometime around 1918 and the red imported fire ant, Solenopsis invicta Buren, in the late 1930's. The presence of imported fire ants in the United States was first reported in 1929 by Loding. Both species probably came to the port in soil used as ballast in cargo ships. In the years preceding the arrival of the red imported fire ant, the black imported fire ant slowly spread into adjacent counties in Alabama and Florida. Since its introduction, the red imported fire ant, a much more aggressive species than the black imported fire ant, has spread quickly. By the time of the first official survey carried out by the USDA in 1953, imported fire ants had invaded 102 counties in 10 states (Culpepper 1953). Today, the red imported fire ant has spread throughout the southeastern United States and Puerto Rico (Fig. 1) replacing the two native species and displacing the black imported fire ant. Currently, S. richteri is found only in extreme northeast Mississippi, northwest Alabama and a few southern counties in Tennessee.
Imported fire ants disperse naturally through mating flights, colony movement or by rafting to new sites during periodic floods. It is through the actions of man, however, that the dramatic spread of imported fire ants has occurred. Beginning soon after the Second World War, and in conjunction with the housing boom of the period, the imported fire ant began its march across the South. The spread of these ants was largely due to the movement of grass sod and woody ornamental plants used in landscaping. This inadvertent movement of S. invicta and S. richteri was noted by the U.S. Department of Agriculture in 1953 when a direct link was established between commercial plant nurseries and the spread of imported fire ants. In response to mounting public pressure, the U.S. Congress appropriated $2.4 million in 1957 for control and eradication efforts. As part of an overall plan a quarantine was imposed to retard or prevent the artificial dissemination of these now notorious pests. On May 6th 1958, regulations governing the movement of nursery stock, grass sod and some other items were instituted through the Federal Quarantine 301.81 (Canter 1981). By that time, however, imported fire ants had moved into eight southern states. This spread, although slowed considerably by federal regulations and climatic conditions, continues even today. In recent years, isolated infestations of imported fire ants have been found as far west as California and as far north as Kansas and Maryland.
Biology
Ants belonging to the genus Solenopsis can readily be distinguished from all other ant species in North America by their 10-segmented antennae with a 2-segmented club. These characteristics, combined with the presence of a sting, a two-segmented pedicel and an unarmed propodeum make identification of the genus relatively easy. Identification of individuals to the species level is somewhat more difficult; made more so by the hybridization between the two "native" species as well as between the two imported species.
Colonies of fire ants consist of eggs, brood, polymorphic workers, winged males, winged females and one or more reproductive queens. Among the sterile workers, labor is divided by age (and to a much lesser degree by size). Younger workers are assigned the job of caring for the developing brood; middle-aged workers are tasked with colony maintenance and protection while the eldest workers forage for food.

Alate, or winged forms, are most abundant in the late spring and early summer but can be found at any time of the year. The winged forms are reproductives. Males, easily distinguished from female alates, are decidedly smaller, glossy black and have a small head (Fig. 2). Although both alate males and females can be found in the same colony; as a general rule one form will be dominant. Nuptial flights most often occur in the middle of the morning one or two days following a rainfall; if the temperature is above 22°C and the wind is light. The males fly first and await the females in the air. The female alates emerge and take flight climbing up into the cloud of waiting males where they mate in the air. After mating, the male dies and the newly mated female lands, sheds her now useless wings and begins searching for a suitable nesting site. The new queen excavates a brood cell approximately 25-50 mm below the surface of the soil. Often more than one new queen will occupy the same cell but only one will survive to establish a colony. Queens do not forage for food but rely on fat reserves and the energy acquired from absorbing her wing muscles to survive until her first workers are ready to take on the task of colony maintenance. Initially, the new queen will oviposit 10-15 eggs. These hatch in 7-10 days and are fed by the queen through trophallaxis (exchange of alimentary fluids) or from sterile, trophic eggs she has laid for that purpose. In 6-10 days, the larvae pupate and emerge 10-15 days later. These workers, called minims, are very small due to the limited amount of energy the queen could devote to their development. These small workers open the brood cell to the outside world and begin foraging for food to feed the queen and the developing brood. Within 30 days, larger workers have emerged and the colony begins to grow. Workers start to emerge daily and within six months several thousand workers can occupy the colony and a "mound" is readily visible. As the colony matures, the polymorphic nature of the worker becomes more apparent. The largest workers in the colony (majors) can be as much as 10 times the size of the smallest workers (media) (Fig. 3). The queen lives up to seven years and produces an average of 1600 eggs per day throughout her life. At maturity, a monogynous fire ant colony can consist of over 250,000 ants.

One of the identifying characteristics of a fire ant colony is the earthen nest or mound. The mound is a conically-shaped dome of excavated soil that has a hard, rain-resistant crust. The mound averages 0.40 m in diameter and 0.25 m in height. In heavier soils, a mound can exceed 1.0 m in height and 1.5 m in diameter. There are usually no external openings in the mound; tunnels approximately 25-50 mm below the surface radiate from the mound allowing foraging workers ready egress and ingress. The purpose of the mound is three-fold: 1) to be a flight platform for nuptial flights; 2) to raise the colony above the water table in saturated ground and; 3) to act as a passive solar collector to supply warmth to the colony during the cold winter months. Although mound size and shape differs to some extent based on soil type, during the dry hot days of late summer and early fall, new mounds are not formed and older mounds are not maintained. While mounds are important to a colony, they are not essential for colony survival. Given a dark, protected site with sufficient moisture and an adequate supply of food, fire ants will nest in a wide variety of sites (e.g. rotten logs, walls of buildings, under sidewalks and roads, in automobiles, in dried cow manure).
Fire ants are omnivorous, feeding on almost any plant or animal material; although insects seem to be their preferred food. In rural habitats, fire ants have a major impact on ground nesting animals from insects to reptiles to birds to mammals. The arrival of imported fire ants into an ecosystem wrecks havoc on the local ecological community. Studies (Allen et al. 1995) have shown that a minimum two-fold reduction occurs among populations of field mice, oviparous snakes, turtles and other vertebrates when imported fire ants are allowed to establish colonies within a given area. In some instances, the depredation by fire ants has completely eliminated some species from an ecosystem (Porter & Savignano 1990). The reduction or elimination of a species or group of species from a system has repercussions throughout the local food web. Not only do imported fire ants reduce animal populations, they also feed on plants. Fire ants attack young saplings and seedlings. They destroy buds and developing fruits and have been shown to feed on the seeds of 139 species of native wildflowers and grasses (Lockley unpubl.). Secondarily, fire ants "nurse" numerous homopteran pests of plants such as aphids and scale insects. Although not conclusively shown, observations indicate that their activity on the plant itself may reduce the ability of pollinators to successfully pollinate flowers.
In agriculture, fire ants have been identified as damaging fifty-seven species of cultivated plants (Adams 1986). Fire ants feed on the germinating seeds of some crops (e.g. corn, sorghum, soybeans) and the buds and developing fruits of others (e.g. citrus, okra). Tunneling fire ants have galleried Russet potato tubers and have significantly damaged the subterranean pods of peanuts. Young citrus and pecan trees have been destroyed when imported fire ants girdled stems and trunks. Damage to plants is exacerbated during periods of drought as fire ants seek alternate water sources. In fields where drip irrigation is used, fire ants will build their mounds over the emitters reducing or blocking the flow of water to crops. In some cases, actual physical destruction of microsprinkler pulsator assemblies has occurred (Boman et al. 1995). Finally, the mere presence of fire ants on plants and within the field will deter hand-harvesting of fruits and vegetables.
As an urban pest, imported fire ants cause many of the same problems experienced in rural areas as well as some problems unique to the urban environment. As in agriculture, imported fire ants cause significant damage to numerous plants and, as in rural habitats, fire ants can reduce the number of birds and mammals in an urban landscape. Fire ants nest within urban structures such as the walls of homes and offices. They establish colonies under sidewalks and roadways. When the site is abandoned, subsidence will cause cracks to appear and will occasionally result in the complete collapse of sections of these structures. The presence of fire ants can deter outdoor activities in yards, parks and school grounds. Home invasions can threaten small children and the elderly. House invasions are especially prevalent during periods of heavy precipitation and flooding. Fire ant colonies have been found inside automobiles, trucks and recreation vehicles (Collins et al. 1993). Traffic accidents have been caused by fire ants stinging the drivers of automobiles. Victims of highway accidents can be attacked by fire ants if they are thrown from their vehicles.
Imported fire ants are attracted by electrical currents and have caused considerable damage to heat pumps, air conditioners, telephone junction boxes, transformers, traffic lights, gasoline pumps, et cetera.
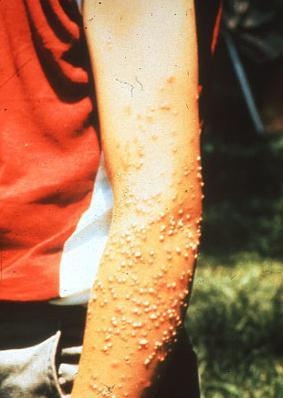
Fire ants are most notorious for their stinging behaviour. They respond rapidly and aggressively to any disturbance of the colony or to a food source. A single fire ant can sting repeatedly (Fig. 5) and will continue to do so even after their venom sac has been depleted. Initially, the sting(s) result in a localized intense burning sensation (hence the name "fire" ant). This is followed within 24 - 48 hours by the formation of a white pustule at the sting site (Fig. 6). This pustule is formed only in response to the stinging of the two imported species. No pustule forms from the stings of native species. These pustules can become sites of secondary infection if not kept clean and can leave permanent scarring. A minority of those stung by fire ants are hypersensitive to the venom and can react quite strongly; suffering chest pains, nausea, dizziness, shock or, in rare cases, lapsing into coma. Some deaths have been documented as having been caused by fire ant stings but these cases are extremely rare.

Control
While any attempts to control imported fire ants over large areas are currently impractical, there are two basic methods that can successfully control fire ants within a limited area: treatment of individual mounds and broadcast treatments.
Individual Mound Treatments
There are several proven methods that can be employed to control individual colonies of fire ants. Insecticidal mound drenches with common insecticides are generally effective against fire ant colonies. The mound is flooded with a large volume of a liquid containing a contact insecticide such as carbaryl, diazinon, dursban, et cetera. Numerous insecticides are currently labeled for this use. A major problem with this method is that the queen is sometimes too deep within the colony to be contacted by the toxicant. Care must be taken not to disturb the mound prior to application of the drench. A disturbance will alert the colony and the queen may be taken deeper into the mound. Application of insecticidal surface dusts or granules have a limited effect on a colony if they are not watered in. The dissolved granules must come into direct contact with the ants to have any effect. As in mound drenches, care must be taken not to disturb the colony prior to application. The queen can be taken to a point within the mound where she may not come in contact with the poison. Some insecticides are marketed as injectants. These are usually more effective than surface applications or mound drenches but are also more expensive and can be dangerous if not handled properly. Fumigants are also commercially available. Again, they are expensive and dangerous if not handled properly. A number of fire ant baits are available. These can be used for treating individual mounds or for broadcast treatment of larger areas. The bait should be uniformly applied around the mound 0.3 to 1.0 meters away and not on the mound itself. Baits are much slower than the control methods listed above but are generally safer, cheaper and more effective in the long run.
Broadcast Treatments
A number of fire ant baits are commercially available for broadcast treatments. Baits are composed of an inert pregelled corn carrier and soybean oil. A toxicant (either a slow-acting insecticide or an insect growth regulator) is incorporated into the oil. Foragers find the bait and carry it back to the colony. Once there, the ants will ingest the soybean oil/toxicant and begin feeding other members of the colony. Eventually, the toxicant is spread throughout the mound and all members of the colony are affected. Broadcasting baits has a number of advantages. Unlike individual mound treatments, colonies need not be "located" in order for them to be treated. Foraging workers quickly retrieve the broadcast granules and carry them back to their nestmates. Broadcasting baits is a great deal less labor intensive than individual treatments and per acre costs are dramatically lower than with any of the other methods. Some of the disadvantages to broadcasting baits include: lack of specificity to fire ants; baits dissolve when they come in contact with water rendering them irretrievable by fire ants; retrieval of the baits is temperature dependent; and baits are slow-acting.
Biological Control
A large number of arthropod species have been identified from nests of imported fire ants. Unfortunately, the vast majority are transients and seem to have no specific relationship with the ants (Collins 1992). However, many of these are known to be endoparasitic, socially parasitic or predaceous (Silviera-Guido et al. 1973, Williams 1980) and studies continue evaluating their effectiveness. Fungal, protozoan and viral pathogens have also been identified in association with fire ants (Jouvenaz 1983). Three potential biological control agents have been identified in South America for intensive study. These studies, if successful, may lead to their introduction into the United States. The organisms under consideration are: Pseudacteon sp. flies (Diptera: Phoridae) ca. 17 spp.; Thelohania solenopsae a protozoan disease and; Solenopsis dagerrei (Hymenoptera: Formicidae), a workerless social parasite.
Phorid flies are known to be parasitic on a number of ant species. Some are thought to be host specific to imported fire ants. The adult flies oviposit on foraging fire ant workers outside the mound; the maggots migrate into the ant's head capsule where they feed. This eventually leads to decapitation of the ant. As interesting as this phenomena is, the major effect of these flies is to cause the ants to cease foraging. In the presence of the fly, worker ants will retreat into the colony to prevent oviposition by the phorid. This disruptive response to the fly restricts the ability of the colony to feed itself and may "even the playing field" so that other ant species can become more competitive with fire ants.
The microsporidian disease Thelohania solenopsae is an obligate intracellular pathogen of fire ants. Preliminary field studies on populations of the black imported fire ant, S. richteri, carried out in Argentina indicated that decreasing densities of fire ants were associated with increasing presence of this pathogen. These data suggest that this pathogen may be an important factor in reducing fire ant numbers by weakening the colonies. Although the vertical transmission of this disease is understood, the horizontal transmission is not.
Solenopsis (Labauchena) dagerrei is a parasitic ant that attaches to the fire ant queen and redirects fire ant workers to tend the brood of the parasite to the detriment of the colony's own larvae. S. dagerrei is intriguing because it lacks a worker caste; only queens and males are produced. The presence of this parasitic ant has a debilitating effect on colony growth and the proportion of sexual reproductives produced in the colony. Queens of S. dagerrei enter fire ant colonies and attach themselves to the mother queen. Previous studies have demonstrated that this parasite inhibits the fire ant queen and her egg production; thus causing the fire ant colony to collapse and eventually die out.
To date, none of these natural enemies has been sufficiently evaluated to determine if, in and of themselves, they might produce any true suppression of fire ant populations. In all likelihood, parasites, predators and pathogens will be used in combinations to reduce colony fitness. Reduction in colony vitality could cause greater mortality under stress conditions and allow for better competition from native ant species.
References
- Adams, C.T. 1986. Agricultural and medical impact of the imported fire ant. IN: C.S. Lofgren & R.K. vander Meer (eds.). Fire Ants and Leaf-cutting Ants. Biology and Management. Westview Press. pp. 4857.
- Allen, C.R., R.S. Lutz, and S. Demarais. 1995. Red imported fire ant impact on Northern Bobwhite populations. Ecol. Appl. 5(3): 632638.
- Boman, B.J., R.C. Bullock and M.L. Parsons. 1995. Ant damage to microsprinkler pulsator assemblies. Appl. Eng. Agric. 11(6): 835837.
- Canter, L.W. 1981. Cooperative imported fire ant programs - Final programmatic impact statement. USDA-APHIS-ADM8101-F. 240 p.
- Collins, H.C. 1992. Control of Imported Fire Ants: a review of current knowledge. USDA-APHIS Technical Bulletin 1807: 27 pp.
- Collins, H.L., T.C. Lockley and D.J. Adams. 1993. Red imported fire ant (Hymenoptera: Formicidae) infestation of motorized vehicles. Fla. Entomol. 76(3): 515516.
- Culpepper, G. H. 1953. The distribution of the imported fire ant in the Southern States. Proceedings of the Association of Agricultural Workers 50: 102.
- Jouvenaz, D.P. 1983. Natural enemies of fire ants. Florida Entomologist 66:111121.
- Loding, H.P. 1929. An ant (Solenopsis saevissima richteri). U.S. Department of Agriculture Insect Pest Survey Bulletin. 9:241.
- Lofgren, C.S. and R.K. Vander Meer (eds.) 1986. Fire Ants and Leaf-cutting Ants: biology and management. Boulder, CO: Westview. 247 pp.
- Porter, S.D. and D.A. Savignano. 1990. Invasion of polygyne fire ants decimates native ants and disrupts arthropod community. Ecology 71(6): 20952106.
- Silviero-Guido, A., J. Carbonell and C. Crisci. 1973. Animals associated with the Solenopsis complex with special reference to Laubauchena daguerri. Proceedings Tall Timbers Conference on Ecology and Animal Control Habitat Management. 4: 4152.
- Williams, R. N. 1980. Insect natural enemies of fire ants in South America with several new records. Proceedings Tall Timbers Conference on Ecology and Animal Control Habitat Management. 7: 123134.