David N. Byrne, Rufus Isaacs and Klaas H. Veenstra
Department of Entomology, University of Arizona
Tucson, Arizona
Introduction
The importance of dispersal in insect population dynamics has been largely ignored when formulating integrated pest management (IPM) programs. This is because studies on dispersal by insect pests have focused largely on long-range migration by moths and locusts at high altitudes. For example, the cases of forecasted dispersal reviewed by Pedgley (1993) all involve migration by insects over considerable distances. Migration on a smaller scale has been somewhat ignored. Some feel that long-range dispersal, which we conveniently define as less than 5.0 km, may be rare (Davis 1980; Loxdale et al. 1993), and more emphasis should be placed on short-range dispersal. Short-range migration is certainly as important biologically as long-range dispersal, and is probably more important agronomically. Laboratory and field experiments concerning migration of agronomic pests on a local scale have largely focused on homopteran insects (Homoptera: Aleyrodidae and Aphididae). Examples are discussed below.
There is no lack of understanding among many academics and growers that migration by pest insects is a phenomenon that impacts crop production. Migration is, however, often omitted as a component of IPM programs at the local level, for two main reasons. To begin, too little is known about the factors that influence migration and dispersal by a particular insect pest. Secondly, testing the impact of this phenomenon on pest populations in agricultural settings is notoriously difficult. This situation is unfortunate, since we require accurate models of insect dispersal to enable realistic forecasting of pest pressure on crops, and this is not possible without detailed knowledge of insect flight behavior and ecology. The interactions between migration and other aspects of pest biology are also critical in developing IPM strategies to deal with mobile pests. For example, in the case of aphids and whiteflies, dispersal of only a few individuals that are insecticide resistant or virus vectors into a crop may give rise to offspring that will cause a significant impact on crop yields.
Definitions
Before discussing specific examples of migratory behavior, we will provide you with important definitions. Migration is a widespread phenomena that has evolved independently in most insect orders (Johnson 1969; Dingle 1972, 1979, 1996; Taylor and Taylor, 1983; Taylor 1986). The definition that best suits the purposes of our research, and that is most consistent with insect behavior we have observed, has been provided by Kennedy (1985).
"Migratory behavior is persistent and straightened-out movement effected by the animal's own locomotory exertions on or by its active embarkation on a vehicle. It depends on some temporary inhibition of station-keeping responses, but promotes their eventual disinhibition and recurrence."
According to Dingle (1996), important characteristics of this definition are:
- persistence; that is, the activity of the insect is tenacious so that it carries the migrant beyond its original habitat to a new one where it gathers previously unavailable resources.
- dispersal is straightened out; that is, there is a directionality to the dispersal and a minimum of turning and backtracking.
- dispersal is undistracted; that is, the insect ignores vegetative cues (cues to resources that could promote growth and maintenance) and keys on other cues, such as sky light, that might be used for activities such as reproduction are employed to support migration.
Flight that has no migratory behavioral component is termed 'trivial flight', and is usually associated with short duration flights between hosts. Crop pest insects that are exhibiting this behavior are attracted to green and yellow wavelengths of light and not to ultraviolet light. In our studies on whitefly flight, insects have been observed in both migratory and trivial flight behavioral phases.
Dispersal is a term that is inclusive of migratory and trivial flight (insects can disperse by either behavior, or by random locomotion).Dispersal can be accomplished by populations and not individuals (Southwood 1981). Dispersal may or may not be associated with the characteristics described above.
Dispersal is more simply defined as movement that results in an increase in the mean distance between individuals.
Both migration and dispersal may lead to emigration of a pest from one crop and its eventual immigration into another. Local dispersal by insects may be effected by migratory behaviors, or by short host-seeking flights.
Local Dispersal by Pest Insects
Aphid Examples
Dispersal by insects on a local scale has been extensively studied in aphids. All of these studies use techniques that show that the insects moved from one place to another, but their behavior during dispersal is not known, i.e., they may or may not be engaged in migratory flight. Thus, we cannot categorically state that the insects were all true migrants in the sense of the definition we have provided. Whatever the behavioral states of the insects, they are primarily relocated to new host plants as a result of being carried by the wind. Some of the aphids that take off and have phototactic flight behavior are evidently carried large distances on the wind (see Loxdale et al. 1993 for review). These insects may embark on windborne migratory flights that can carry them for great distances, but the ecological importance of this has recently been challenged (Loxdale et al. 1993). They argue that while long distance dispersal is possible, the majority move locally.
Evidence for local migration by pest insects has been collected from a number of reports on the green peach aphid, Myzus persicae (Sulzer). For example, a variant of this species (R2) that is particularly resistant to insecticides was found to be concentrated in northern Britain in samples taken between 1980 and 1984, and the incidence of resistant aphids had not changed appreciably since surveys in 1976 (Furk 1986). Other resistant variants of this species have been surveyed, and are known to have stable regional distributions (ffrench-Constant and Devonshire 1988). This situation would be predicted by a local dispersal of individuals with limited regional gene flow. In the UK, green peach aphid populations in the south and west are more susceptible to insecticides than those in the north, and thus it seems that there is limited interaction between these populations.
As discussed in Loxdale et al. (1993), we must have knowledge of insect population dispersal to be able to understand dispersal of viruliferous or resistant pests and forecasting pest outbreaks. This is especially true if we need to predict these occurrences or design management programs that take dispersal into account. The control of virus disease in Idaho in the 1960's is a good example of this approach, demonstrating how short-range flight of insects can be economically important.

In the early 1960's, potato crops in Idaho, United States (US) were suffering large economic losses to viruses, especially to potato leaf roll virus that is transmitted by green peach aphids and causes stunted growth and yellowing of foliage. A strong correlation was found between the number of aphids per 100 leaves in the potato crop, and the fieldís distance from principal towns. In addition, within field sampling showed a greater number of aphids, and higher disease incidence on the side of the field closest to the towns (Bishop 1965). Viruliferous aphids and potatoes were found in the towns' gardens. Problems were related to the fact that is was common practice for gardeners to use small, diseased potatoes as seed pieces. Short duration flights of aphids from the gardens to the surrounding fields were implicated as the cause of the disease outbreaks. Based on this implied local migration by green peach aphids, a control program was implemented in an area-wide program in southeastern Idaho beginning in 1961 (Bishop 1967). There were three main components to this IPM program: 1: supplying seed potatoes certified as being virus free to home gardeners, 2: malathion application to transplants and gardeners' potatoes and 3: removal or treatment of peach and apricot seedlings with oxydemeton-methyl where the insects overwinter. Implementation of this program drastically reduced aphid numbers in the potato crop to less than 1% of the pre-program level, and significantly reduced the incidence of the virus.
Whitefly Examples
Introduction

The focus of our laboratory's research is the ecology of the sweet potato whitefly, Bemisia tabaci (Gennadius). This insect has been observed in the US since the early part of this century (Russell 1975). It has the ability to transmit a number of viral pathogens (Brown and Nelson 1986) and additionally reduces yield through direct feeding damage (Perring et al. 1991, Johnson et al. 1992, Riley and Palumbo 1995 and Palumbo and Sanchez 1995). More recently, the problems associated with honeydew (whitefly excreta) and sooty mold, Capnodium spp., (i.e.,stickiness and staining) have become more important for growers across the southern tier of the US and around the world (Hector and Hodkinson 1989; Byrne and Miller 1990).
Although sampling plans and action thresholds are now being developed for melons, (Tonhasca et al. 1994a and 1994b; Palumbo et al. 1996) and cotton, Gossypium spp. (Dennehy et al. 1995), only a few current management programs incorporate more than a few of the strategies we associate with IPM. Growers of most crops are primarily left with one principal option, i.e., the direct application of pesticides. The situation mandates the need for alternative control strategies. We are convinced that a better understanding of flight behavior by Bemisia will lead to the development of new cultural control techniques involving computer simulation models that can predict when, where and how far Bemisia isgoing to disperse.
Initial Experiments
An early concern, which was pivotal, was whether B. tabaci was capable of exhibiting migratory behavior. We had discovered that wingbeat frequency was not significantly correlated with wing loading for these small insects (Byrne et al. 1988). Larger insects seem to employ a strategy similar to other flying animals, i.e., they compensate for high wing loading with higher wingbeat frequencies. The lack of correlation for these two values in whiteflies led to erroneous assumptions concerning the ability of whiteflies to sustain flight. On the other hand, we knew that the internal lipids in adult females of B. tabaci represented 40% of their total dry weight(Cohen and Byrne 1992). These compounds are commonly thought to serve as an energy source for insects during flight. For comparison, in the pea aphid, Acrythosiphum pisum (Harris), which is known for its migratory ability, lipids account for only 10% of the dry weight. These findings suggested that, despite their small size (33 mg [Byrne et al., 1988]), B. tabaci might possess enough energy stores to engage in extended flight.
Laboratory Behavioral Studies
To investigate a series of hypotheses, a vertical flight chamber (Blackmer and Phelan 1991 was used to examine untethered flight of& B. tabaci. Blackmer and Byrne (1993a) characterized the periodicity, intensity and duration of whitefly flight. Behaviorally they reaffirmed that there was a dichotomy between groups of whiteflies relative to flight behavior. Some flew for more than 3 h, others flew sporadically, some flew not at all.
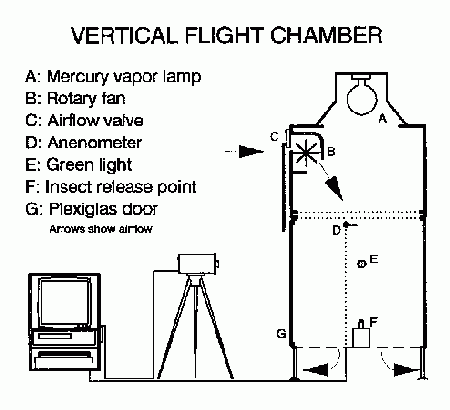
Blackmer and Byrne (1993a) also examined how whitefly age and host quality (vegetative vs. senescing) affected flight activity. The propensity to takeoff, proportion exhibiting phototactic orientation, and flight duration varied with whitefly age. Longer-duration flights occurred between three and five days of age. Six days following eclosion little flight took place because, among other things, indirect flight muscles and mitochondria in the thorax had started to deteriorate (Blackmer et al. 1995). Host quality also influenced the timing of flight behavior. Whiteflies reared on senescing plants exhibited greater takeoff rates and initiated longer phototactic flights than individuals reared on vegetative plants.
Laboratory Physiological Studies
It is often assumed that there is a trade-off between migratory capability and reproduction. In migrating individuals, nutritional and energy resources are diverted away from growth and reproduction towards building and maintaining flight muscles and energy resources needed for flight. This allocation of resources into either migration or reproduction has been called the "oogenesis-flight syndrome" (Johnson 1969), and has been well documented in alate aphids. We found small, but significant increases, in egg-protein levels were found with longer flights and increasing weights. This was in contrast to the expected (Blackmer et al. 1995).
Lipids are the most suitable materials for storage of energy reserves. Compared to carbohydrates, lipids can supply as much as eight times more energy per unit weight (Beenakkers et al. 1985). In addition, twice as much metabolic water is produced by metabolizing lipids instead of carbohydrates (Downer 1985). A high lipid content associated with migration has been found in many aphids and some bark beetles. In the sweet potato whitefly we found no evidence that lipids are stored for dispersal of less than 3 km. However, flight chamber experiments indicate that in females, lipid content is correlated with flight duration (unpublished data). Thus, lipid reserves could be of more importance for long distance migration.
Understanding the factors leading to production of migratory morphs is important for IPM strategies. Physiological studies can lead to valuable insight into migratory capabilities and will allow us to estimate the effective range of dispersal of insect pests.
Field studies with whiteflies
In order to corroborate laboratory observations, field experiments were conducted beginning in 1992 (Byrne et al. 1996). Initial experiments concentrated on the impact of wind flow on the directionality of flight by B. tabaci. A 0.73 ha circular field was planted with cantaloupe, Cucumis melo L. The area around the field was surveyed for placement of muffin fan traps along a series of 16 transects placed 22.50 apart. These traps are described in Byrne et al. (1996). They allowed the capture of living whiteflies.
The evening before each sampling day the melon field was dusted with fluorescent dust. Tracking insects from where they alight to their landing place is possible in the field, and has been done with small insects with some success using the techniques listed separately. The traps were turned on the morning of each sampling day (at approximately 6:00 a.m.). After 3 h, whiteflies marked with dust were collected from the traps. For each sampling date in 1992 the relationship between wind flow and the number of whiteflies captured in traps in the direction opposite the prevailing wind was evaluated. Prevailing wind directions from the northeast (Brown et al. 1995) and trap catch were in agreement for all dates (when the wind was from the northeast > 90% of the whiteflies were caught in traps placed to the southwest of the marked field). This provided empirical evidence that whitefly migration is wind-aided.
Following 1992, traps were located as part of a grid to the southwest of the marked field. Maps of fields and trap locations were digitized using a digitizing tablet. Data were analyzed using the Geostatistical Information System (GIS). The principal feature evident when we examined the data from our field experiments was that whitefly distribution following dispersal was patchy. Almost all of the sampling dates show greatest trap catches of marked insects near the marked field and on the periphery of the grid than in the center of the grid. Pooling data from all sampling dates in 1993 produced a line graph indicating a bimodal distribution. It is evident that the trap catches did not indicate a simple diffusion pattern, a pattern that might be expected if dispersal was passive across featureless topography. This is in contrast to patterns seen by Turchin and Thoeny (1993) who described dispersal by southern pine beetles, Dendroctonus frontalis Zimm., using a diffusion model. We believe our pattern may be explained by the existence in whitefly populations of two behaviorally distinct types; trivial flying individuals and migratory individuals as described by Byrne and Houck (1990) and Blackmer and Byrne (1993 a and b). Insects caught in the immediate vicinity of the source field are likely to be trivial fliers, while those caught on the grid perimeter are thought to be migratory individuals. The explanation for the relative paucity of whiteflies caught in the middle of the grid is that migratory whiteflies do not fly towards green vegetation until they have flown for a given period of time (Blackmer and Byrne 1993a).
In our studies, the greatest distance marked whiteflies have been trapped is 2.7 km from a source field (Byrne et al. 1996). Since whiteflies were consistently captured in the most distant traps, the effective migrational range of B. tabaci may be considerably greater than this. Future experiments will be conducted with an expanded grid to further define the effective migrational range (i.e., the distance they can travel and still initiate a colony) of B. tabaci.
Field research is still ongoing, and these experiments have already revealed a few surprises. For example, we have found that whiteflies were caught in traps placed at the edge of the source field, at over 20 ft above the crop. Traps were in place just before dawn, because the air starts to move at sunrise, and flight is observed as soon as there is sufficient light available to stimulate take-off. Thus, it would seem that the migrants are taking maximum advantage of the low wind speeds to gain height and take them away from the plant boundary layer. This has far-reaching implications for our understanding of whitefly migration and their dispersal on the wind. That whiteflies can take off and ignore plant cues in the field also reinforces our faith that the behaviors observed in the laboratory are those employed by the insects in the field.
Development of predictive models
Ideally, our knowledge of insect migration would lead to the development of simulation models, that serve as an aid to growers in making management decisions. To our knowledge, no such models exist as of yet. This is, in our opinion, partially due to some inherent difficulties in developing and implementing such models.
The "art" of modeling involves reducing a complex biological reality down to a few mathematical statements. For example, one could treat a field of melons, as one unit, with specified size, location, and time of planting. However, the development of whiteflies in a given melon field and their migratory tendencies, are likely to be influenced by the many other factors, like watering regime, amount of fertilizer applied, soil type, or disease incidence. In order to incorporate these factors into the model we need to know the effect of each of these factors on whitefly biology. If one of these factors turned out to be critical for the model, growers would have to spend time (and money) collecting data on this variable. That would severely diminish the value of these models as a management tool.
In addition, one typically has only data available from a small number of successive years for the development of migration models. Thus, the model will be based on a subset of potential environmental conditions. For example, the years for which data were available could be characterized by relatively cool summers. Whether the model thus developed could be extended to years with different temperature regimes can only be examined by collecting data over longer time periods.
Summary
The migratory behavior of pest insects is an important aspect of their life cycle that has obvious implications for where populations will be exerting pressure on crop production. Our research on whitefly flight behavior has already yielded information that can be used to assist growers when making decisions concerning crop placement and planting times. The important findings with relevance to IPM are summarized below:
- Small insect pests can have the capacity for extended flights, controlled by migratory behavior.
- Insect dispersal is often in response to both biotic and abiotic conditions that promote migratory behavior.
- Migration by insects is an adaptation to a rapidly changing environment. The way in which biotic and abiotic factors influence migration differs widely between species.
- Wind speed and direction play a significant role in dispersal of small insects.
- In many insects, migrating individuals are different from non-migrating individuals in physiology and/or morphology.
- Dispersal by insects acquires additional importance for IPM when they are carrying genes for resistance or are vectors of plant pathogens.
- They can disperse large distances, but most whiteflies land less than 5.0 km from their original point of take-off.
- Crop placement and timing should take account of the likely pest pressure from mobile insects.
- Models can be constructed that help in predicting insect dispersal, but they are only as good as the information they are based on.
- Knowledge of migration is important for many IPM programs. This does require, however, detailed knowledge of the migratory capacity of each pest insect species.